La seconda parte di questa discussione è stata elaborata della CIRED dell’Università CaFoscari di Venezia e quì riportata a vantaggio di quegli studenti che vogliono approfondire più accuratamente l’argomento.
Una pianta per vivere ha bisogno di nutrirsi esattamente come tutti gli altri esseri viventi,ma al contrario delle piante, gli altri esseri viventi assumono il nutrimento dall’esterno (ETEROTROFI).
Le piante, invece,«si nutrono da sole», cioè fabbricano il loro nutrimento all’interno della stessa pianta (AUTOTROFE) prelevando dall’esterno sostanze non organiche trasformandole in organiche.
Ci poniamo a questo punto una domanda:
COME FA LA PIANTA A PROCURARSI LE SOSTANZE CHE UTILIZZA PER RICAVARE L’ ENERGIA CHE LE SERVE PER MANTENERSI IN VITA E PER COSTRUIRE LE VARIE PARTI DELLA PIANTA ?
Attraverso le radici, la pianta assorbe dal terreno acqua e sostanze nutrienti mentre attraverso le foglie assorbe l’anidride carbonica dell’aria ed e’ proprio nella foglia che H2O e CO2 si uniscono per formare il glucosio (C6H12O6) che e’ l’elemento chiave per il funzionamento generale della pianta.Pertanto la foglia rappresenta un vero e proprio laboratorio chimico importantissimo, quindi è necessario conoscere come e da che cosa è formata una foglia e conoscere le funzioni delle varie parti di essa.
Per cominciare, diciamo che la foglia è un insieme di cellule collegate tra loro contenente all’interno nucleo, citoplasma ed organelli cellulari vari. Al contrario delle cellule animali, le membrane cellulari di una pianta sono rigide ed il citoplasma contiene degli organelli specifici contenenti clorofilla e carotenoidi in quantità preponderante ma ha in comune con le cellule animali i ribosomi dove si fabbricano le proteine ed mitocondri dove si produce ATP adenosin trifosfato.
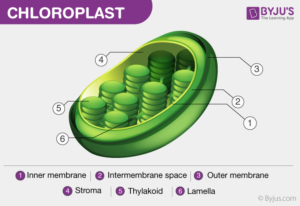
Gli organelli cellulari a ridosso della parete sono i cloroplasti che contengono le clorofille ed i carotenoidi. Un cloroplasto e la sua sezione vista al microscopio, è rappresentato nella figura seguente :
/cutaway-illustration-of-plant-chloroplast-680790343-588b47f83df78caebcfceb00.jpg)
Presenta due membrane cellulari una esterna ed una interna. Vi sono quindi dei dischi chiamati tilacoidi che formano delle pile ognuna delle quali è chiamata Grano e questi sono collegati tra loro. All’interno delle membrane dei tilacoidi vi sono due sistemi che catturano la luce solare e la trasformano in composti chimici ad alta energia che è utilizzata poi nello stroma cioè lo spazio all’esterno dei tilacoidi, per fabbricare il glucosio.
Ogni fotosistema contiene da 250 a 400 molecole di pigmenti e numerose proteine.
un fotosistema è costituito da
COMPLESSO ANTENNA:
costituito da Clorofille e carotenoidi.
Questo sistema raccoglie l’energia luminosa e la convoglia al centro di reazione. L’energia acquisita dalla luce si trasmette alle molecole per risonanza.
CENTRO DI REAZIONE
In grado di convertire l’energia luminosa in energia chimica. In questo caso si ha anche trasferimento di elettroni.




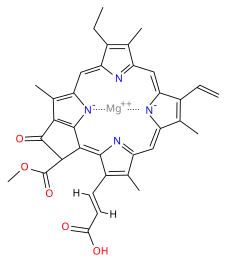

queste sono le molecole di clorofilla a, b, c1 e c2 in cui al centro degli anelli pirrolici vi è il Magnesio.
Clorofilla a:
è presente in tutti gli organismi con Fotosintesi che produce ossigeno
Clorofilla b:
caratteristica degli organismi fotosintetici verdi (piante vascolari, briofite, alghe verdi, Euglenoidi).
Clorofille c1,c2:
pigmenti accessori in diversi gruppi Diatomee, Feofite (alghe).
Batterioclorofille:
presenti nei batteri con fotosintesi che non produce ossigeno
I pigmenti principali delle piante superiori sono le clorofille ed i carotenoidi.
La clorofilla a è il primo attore della fotosintesi in tutte le piante; la clorofilla b e gli altri pigmenti sono “pigmenti accessori” o “pigmenti antenna“, che raccolgono una parte dell’energia luminosa e la trasmettono alla clorofilla a.
La parte che conta per l’assorbimento della luce è l’anello porfirinico al centro del quale vi è un atomo di magnesio. L’anello porfirinico possiede, infatti, un gran numero di doppi legami coniugati ed è costituito essenzialmente da 4 eterocicli (anelli contenenti anche atomi diversi da C) collegati da ponti CH.
L’energia luminosa viene catturata dagli elettroni dell’anello porfirinico che passano dal loro livello energetico fondamentale ad uno stato eccitato. Le clorofille sono dei fotorecettori molto efficienti.
Quando una delle molecole di clorofilla viene eccitata da un fotone hν la molecola eccitata trasferisce, per risonanza induttiva, la sua eccitazione ad un’altra clorofilla adiacente e così via, finché quella eccitata che è adiacente ad una clorofilla che si trova nel centro di reazione (che è ad energia più bassa), trasferisce a quest’ultima la sua energia.
La risonanza induttiva è un processo per cui una molecola eccitata può trasferire la sua energia ad un’altra molecola adiacente, che passa, a sua volta, ad uno stato eccitato.
Essa può ricevere l’eccitazione da tutte le altre molecole dell’unità fotosintetica, ma non può ritrasmetterla.
Riassumendo il principale carattere comune dei pigmenti fotosintetici come la clorofilla è la presenza di una serie di legami doppi e singoli alternati, detti legami doppi coniugati e quanto maggiore è il numero di legami doppi coniugati in una molecola, tanto maggiore è la lunghezza d’onda assorbita (perciò tanto minore l’energia della radiazione).
I pigmenti fotosintetici dei cloroplasti non sono mai liberi in soluzione: sono ancorati a membrane e organizzati in una serie di complessi dei pigmenti Antenna.
gli spettri di assorbimento della clorofilla a e b sono diversi; questi due tipi di clorofilla si completano l’un l’altro nell’assorbimento della luce solare incidente.
Le molecole di clorofilla hanno un anello porfirinico idrofilo e una catena idrofoba.
L’anello porfirinico è costituito da 4 eterocicli collegati da ponti CH e con i 4 N rivolti verso l’interno che coordinano lo ione metallico al centro.
Qui è rappresentata la clorofilla a.
La clorofilla b è di poco diversa: ha un formile (-CHO) al posto di un metile (-CH3). (in alto a dstra)
Alla catena di trasporto partecipano anche gli atomi di N.
Gli stati eccitati della clorofilla
Lo stato eccitato della clorofilla ad energia più alta corrisponde alla lunghezza d’onda minore (430 nm); quello ad energia più bassa (670 nm) è quello utilizzato nella fotosintesi.
Il passaggio dallo stato eccitato più alto a quello fondamentale (decadimento) non comporta emissione di fotoni luminosi, poiché il salto energetico corrisponde alla zona dell’infrarosso: l’energia viene cioè dispersa sotto forma di calore.
Spettro di assorbimento
E’ il diagramma che mostra, qualitativamente e quantitativamente, le lunghezze d’onda assorbite da una sostanza, in questo caso dal pigmento.

In ascissa la lunghezza d’onda λ , (espressa in nanometri nm), in ordinata l’entità dell’assorbimento o assorbanza, che può venire espressa in varie unità di misura (qui come coefficiente molare di assorbimento o di estinzione che si indica con ε).Lo spettro di assorbimento delle clorofille (la a in azzurro , la b in rosso) presenta due picchi diversi nel rosso e nell’azzurro, separati da un profondo avvallamento (500-600 nm) in cui l’assorbimento è basso. In questo avvallamento, sono più attivi i carotenoidi che così completano l’assorbimento delle radiazioni luminose nella pianta.
La misurazione dello spettro di assorbimento, ovvero del colore, della clorofilla viene svolta con metodiche di spettrofotometria
La misura dell’assorbanza della luce è complicata dal solvente utilizzato per estrarre la clorofilla dalla materia vegetale, che influenza i risultati ottenuti;
- Nel dietil etere, la clorofilla a presenta i massimi di assorbimento approssimati a 430 nm e 662 nm, mentre la clorofilla b li presenta a 453 nm e 642 nm.
- I picchi di assorbimento della clorofilla a sono a 665 nm e 465 nm. La clorofilla a diviene fluorescente a 673 nm. Il picco del coefficiente di assorbimento molare della clorofilla a supera 105 M−1 cm−1, che è il più alto tra i composti organici.
Il fatto che la clorofilla a e b abbiano dei spettri di assorbimento differenti si traduce in un migliore assorbimento della radiazione solare.
Nella tabella di seguito vengono indicate le lunghezze d’onda in nanometri a cui corrisponde il massimo dei picchi di assorbimento primari e l’estensione dei picchi (indicata tra parentesi):
| Clorofilla a | Clorofilla b | Clorofilla c | Clorofilla d | Clorofilla e | |
|---|---|---|---|---|---|
| Picco 1 | 430 nm | 480 nm | 434 nm | (circa 400-470 nm) | – |
| Picco 2 | 663 nm | 650 nm | 666 nm | 700 nm (700-730 nm) | (715-725 nm) |
Carotenoidi
I carotenoidi sono pigmenti di colore giallo o arancione la cui molecola ha due anelli esagonali (aperti o chiusi) collegati fra loro da una lunga catena isoprenica con doppi legami coniugati.Sono idrofobi , essendo generalmente costituiti esclusivamente da C ed H
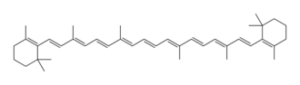
Il loro spettro di assorbimento ha un unico massimo sull’azzurro. Oltre a quella di pigmenti accessori i carotenoidi hanno un’altra importante funzione: proteggono la clorofilla dalla fotoossidazione, una reazione tra clorofilla e ossigeno atmosferico, attivata dalla luce.
Complesso di pigmenti antenna
I pigmenti fotosintetici, clorofille e carotenoidi, sono a stretto contatto tra loro essendo ancorati, data la loro natura liposolubile, allo strato lipidico della membrana.
La cattura della luce è efficiente per la cooperazione di alcune centinaia di molecole di pigmenti, che funzionano come “antenne” per catturare la luce.l complesso trasferisce poi l’energia al centro di reazi0ne. Quindi la maggior parte delle molecole di pigmento possono assorbire radiazioni, ma soltanto alcune molecole di clorofilla, quelle presenti nei centri di reazione, sono in grado di indurre le trasformazioni della luce in energia chimica.
Il centro di reazione contiene una coppia di molecole di clorofilla a circondate da gruppi chimici in grado di accettare l’elettrone eccitato.
In un certo senso il segreto della fotosintesi consiste nella capacità di queste molecole di clorofilla di produrre elettroni ad alta energia per assorbimento della luce e di trasferirli ad un’altra molecola mediante una reazione redox.
Le reazioni fotochimiche avvengono sfruttando la reattività di una molecola allo stato eccitato che, anziché perdere energia attraverso l’emissione di un fotone di fluorescenza, trasferisce un elettrone ad un accettore.
L’elettrone perduto viene poi recuperato a spese di un donatore di elettroni nella fotosintesi dell’acqua.
Trasporto degli elettroni fotosintetici
In questa fase l’energia viene utilizzata per far funzionare la catena di trasporto degli elettroni fotosintetici ad alta energia prodotti nei centri di reazione, attraverso una serie di trasportatori elettronici.
Nella fotosintesi che produce ossigeno è l’acqua il donatore di elettroni; ma data la sua stabilità, è necessario un apparato complesso che catturi l’energia.
Per questo si è evoluto un meccanismo che utilizza due trasduttori accoppiati, chiamati
FOTOSISTEMA I E FOTOSISTEMA II
I due fotosistemi, in coppia, riescono a trasferire elettroni dall’acqua al coenzima NADP+ che viene così ridotto a NADPH.
Il flusso di elettroni attraverso i fotosistemi genera un gradiente protonico che favorisce la sintesi di ATP (FOTOFOSFORILAZIONE)
Praticamente il passaggio di elettroni dall’acqua a NADP porta alla formazione di NADPH, ATP e ossigeno.
I Fotosistemi I e II sono due complicate strutture molecolari che attraversano il doppio strato lipidico delle membrane tilacoidi e sono formate dai complessi di pigmenti antenna, combinati con le clorofille dei centri di reazione e con alcuni trasportatori di elettroni.
Nel fotosistema I il complesso di molecole antenna è costituito principalmente da clorofilla a.
Nel fotosistema II è abbondante la clorofilla b, ma anche in questo caso il centro di reazione è costituito da clorofilla a.
Il fotosistema I ha il massimo di assorbimento a lunghezza d’onda 700 nm, perciò viene chiamato P 700 (P = pigmento); per lo stesso motivo il fotosistema II, è chiamato P 680.
I fotosistemi agiscono come piccoli condensatori elettrici, accumulando energia mediante la separazione e la stabilizzazione delle cariche positive e negative sulle due facce separate della membrana tilacoide.
L’interazione dei due sistemi è mostrata in figura e rappresenta le azioni ossidoriduttive dei due fotosistemi sotto l’azione delle radiazioni luminose assorbite.


FAD = flavin adenin dinucleotide FeS = ferro solfo proteina PQ = plastochinone PC = plastocianina Fd = ferredossina
Esaminando i potenziali standard di ossidoriduzione riportati nello schema a Z si può constatare che il fotosistema II (che estrae elettroni dall’acqua e riduce il fotosistema I) si comporta come un forte ossidante e un debole riducente.
Il fotosistema I, che è in grado di cedere elettroni a un potenziale molto più negativo rispetto al fotosistema II, genera un riducente così forte da ridurre NADP+, comportandosi da forte riducente e blando ossidante.
NADPH è un riducente adatto alle riduzioni biosintetiche, prima fra tutte quella dell’anidride carbonica a zucchero.
Fotosistema II
Nel fotosistema II (chiamato anche PS II, da Photo System II), formato da proteine e pigmenti, sono presenti, fra i trasportatori di elettroni, la feofitina e i plastochinoni.
Il complesso che cattura la luce (LHC) contiene circa 200 molecole di clorofilla a e b legate ad alcune catene polipeptidiche: il gruppo del centro di reazione contiene altre 50 molecole di clorofilla a. All’interno è localizzato un complesso molecolare enzimatico per l’ossidazione dell’acqua; infatti trasferisce gli elettroni dall’acqua ad un chinone, il plastochinone, liberando contemporaneamente ossigeno
Gli elettroni ricchi di energia, dopo essere passati attraverso una serie di trasportatori simili a quelli delle membrane mitocondriali, raggiungono i centri di reazione del fotosistema I.
Fotosistema I
Il fotosistema I (chiamato anche PS I, da Photo System I) genera un forte riducente che porta alla formazione di NADPH che entrerà poi nel ciclo di riduzione dell’anidride carbonica.
Il trasporto di elettroni dall’acqua direttamente a NADPH può essere rappresentato con lo schema a Z di Hill-Bendall dove le variazioni energetiche e l’andamento del flusso degli elettroni dall’acqua all’ NADPH sono meglio descritte in termini di potenziale di ossidoriduzione.
Reazione globale del fotosistema I

Lo schema rappresenta, in sintesi, la reazione globale che avviene a carico del fotosistema I.
PC rappresenta la plastocianina P700 rappresenta il centro di reazione
A0 e A1 rappresentano molecole di clorofilla accettori di elettroni
Ferridossina è la forma ossidata della ferrodossina.
Reazioni del fotosistema I
Nel fotosistema I la luce viene convogliata da un complesso di pigmenti antenna al centro di reazione P700.
Un elettrone viene trasferito dal P700* (stato eccitato della clorofilla) ad una clorofilla accettrice, chiamata A0, formando A0– e P700+. A0– è un riducente molto forte (Eo = -1,1 V), praticamente la biomolecola più riducente.
P700+ cattura un elettrone dalla plastocianina ridotta, ridiventando P700, disponibile per una successiva reazione.
Da A0 gli elettroni vengono trasferiti ad un secondo accettore A1 (che pare essere una molecola di vitamina K) e poi ad una serie di centri ferro-zolfo (ferredossine) all’interno del fotosistema I. La tappa finale è la riduzione della ferridossina, una proteina idrosolubile contenente un centro 2Fe-2S, a ferrodossina.
Questa reazione avviene sulla faccia della membrana tilacoide rivolta verso lo stroma.
Gli elettroni ad alto potenziale della ferrodossina sono poi trasferiti a NADPH+ per formare NADPH
Reazione globale del fotosistema II

Ph rappresenta la feofitina. PQ(QA) e PQ(QB) il plastochinone nei due siti. PQH2 la forma due volte ridotta del plastochinone. b6f il complesso del citocromo PC la plastocianina Z un intermedio che permette di utilizzare acqua come riducente.
Reazioni del fotosistema II
Nel centro di reazione P680 l’energia luminosa viene convertita in separazione di cariche. P680* (centro di reazione eccitato) trasferisce, in pochi picosecondi, un elettrone alla feofitina (Ph), producendo P680+ e Ph–. Questo processo consuma buona parte dell’energia del fotone assorbito.
L’elettrone passa dalla feofitina ridotta al plastochinone legato ad un sito proteico QA (sempre ancorato al fotosistema) e poi ad un altro plastochinone su un sito QB, con l’intervento anche di atomi Mn e ioni Cl– e Ca++.
QB, acquistati due elettroni e due protoni in due cicli di assorbimento di fotoni, esce dal complesso del fotosistema II ed è sostituito da un QB non ridotto. QB ridotto (o PQH2) è riossidato dal citocromo b6f (plastochinolo-plastocianina ossidoriduttasi) e questo viene a sua volta riossidato da una piccola proteina, la plastocianina PC.
Questi processi portano ad un gradiente protonico attraverso la membrana e alla riduzione della PC (che trasporta poi elettroni al fotosistema I). L’energia viene usata per la sintesi di ATP(fotofosforilazione non ciclica).
Il radicale cationico P680*+, formato dal fotosistema II nella tappa della separazione di carica, è un forte ossidante e, mediante un intermedio Z, sottrae elettroni all’acqua generando O2
Feofitina

La feofitina è una porfirina identica alla clorofilla a, salvo per il fatto che è priva di magnesio.La linea spezzata che esce in basso a destra di ognuna delle due molecole rappresenta una catena alifatica idrofoba.
Ossidazione dell’acqua nella fotosintesi
La proprietà più significativa del fotosistema II è quella di usare l’acqua come donatore di elettroni La reazione è:
2 H2O → O2 + 4 H+ + 4 e
Il cuore del sistema per l’ossidazione dell’acqua è un costituente del fotosistema II, un complesso contenente 4 ioni manganese nel suo sito catalitico. Questo complesso viene ossidato dal P680+ in una sequenza di cinque stadi di ossidazione diversi.Gli ioni manganese sono fondamentali nel processo.
Orologio di ossidazione dell’acqua

è un meccanismo ciclico che porta elettroni alle clorofille del fotosistema II.
Ogni fotone assorbito da P680 fa avanzare l’orologio di un gradino negli stati di ossidazione, rilasciando un elettrone.
Quando viene raggiunto l’ultimo stato di ossidazione (IV), l’orologio produce O2 e ritorna allo stato iniziale (X).
Gli stati di ossidazione sono generalmente indicati dalla sequenza:
S0 (X) == S1 (I) == S2 (II) == S3 (III) == S4 (IV)
Flusso di elettroni nella fotosintesi
Semplificando moltissimo, il trasporto degli elettroni nella fase luminosa può essere rappresentato come in figura, secondo la reazione di ossidoriduzione:
2 NADP+ + 2 H2O + hν → 2 NADPH + 2 H+ + O2
In condizioni normali, un elettrone non potrebbe abbandonare l’acqua per ridurre NADP+ poiché la reazione è energeticamente sfavorita.Questo trasferimento di elettroni avviene infatti sfruttando l’energia dei fotoni assorbiti dal fotosistema II.
Per rendere possibile la reazione bisogna coprire un intervallo energetico che va da 0,82 V della coppia H2O/O2 a -0,32 V della coppia NADPH/NADP+
L’energia richiesta nella fotosintesi viene fornita dalla collaborazione dei due centri di reazione dei fotosistemi che operano in serie, riuscendo così a trasferire gli elettroni dall’acqua a NADP+
Fotofosforilazione
E’ un processo che corrisponde alla sintesi di ATP provocata dalla luce.
La sintesi di ATP è legata al flusso di elettroni, che può essere ciclico o non ciclico; ai due tipi di flusso corrispondono la fotofosforilazione ciclica e non ciclica.
Nel flusso ciclico il movimento avviene lungo un percorso che si chiude ad anello con il P700 del fotosistema I: gli elettroni passano dall’accettore primario del fotosistema I a un trasportatore che non fa parte della via Z, il citocromo b6.
Nel flusso non ciclico il movimento degli elettroni avviene lungo l’intera via Z perché gli elettroni iniziano e terminano il loro spostamento a livello di sostanze diverse.
Il meccanismo che accoppia la sintesi di ATP con il trasporto degli elettroni nei cloroplasti viene spiegato dall’ ipotesi chemiosmotica di Mitchell: la produzione di ATP è accoppiata alla formazione di un gradiente di protoni, a sua volta dovuta al particolare orientamento dei complessi molecolari della membrana tilacoide (asimmetria nella membrana tilacoide).
Fotofosforilazione e sintesi di ATP
Nelle membrane tilacoidi c’è un sistema molecolare, comprendente un complesso proteinico detto CF1, a destra nel disegno, che accoppia il gradiente di protoni alla sintesi di ATP.

Il meccanismo esatto del suo funzionamento, sicuramente molto complesso, non è ancora perfettamente chiarito.
Ipotesi chemioosmotica di Mitchell
Nella fosforilazione ossidativa la sintesi di ATP è accoppiata al flusso di elettroni da NADH e FADH2 all’ossigeno, mediante un gradiente di protoni che si instaura tra le due facce della membrana interna mitocondriale.
Si creano così zone a diverso PH come indicato appunto nella ipotesi chemiosmotica che prevede il coinvolgimento di sistemi enzimatici con spesa di energia.
Alcuni componenti della catena respiratoria, NAD e FAD, trasportano sia elettroni che protoni, altri, come i citocromi, trasportano solo elettroni.
Quando gli elettroni giungono ai citocromi, i protoni vengono scaricati nel compartimento esterno, grazie al preciso orientamento dei trasportatori di elettroni nello spessore delle criste mitocondriali e all’energia che si libera durante il flusso di elettroni.
L’energia spesa viene poi recuperata mediante la sintesi di ATP nella ATP sintetasi e questo avviene in varie fasi dei processi di trasporto degli elettroni e fosforilazione ossidativa
Trasporto degli elettroni e fosforilazione ossidativa

viene mostrato lo schema del percorso degli elettroni e la formazione di ATP come prodotto del trasporto elettronico lungo la catena.
Si formano 3 molecole di ATP per ogni molecola di NADH ossidata.
Dall’ossidazione di una molecola di FADH2 si formano invece solo 2 molecole di ATP in quanto i suoi elettroni entrano nella catena respiratoria a livello dell’Ubichinone Coenzima Q saltando il primo complesso che trasporta protoni.
Nei cloroplasti, la zona apH più basso (pH 5) è all’interno della membrana tilacoidale,
La sintesi di ATP, favorita dalla retrodiffusione dei protoni attraverso le ATPsintetasi, dalla zona a pH minore a quella a pH maggiore, avviene comunque nella zona a pH maggiore, perciò entro lo stroma nel caso dei cloroplasti,
Gradiente di protoni nella fotosintesi
Si suppone che i primi protoni del gradiente derivino dalla scissione dell’acqua sulla superficie della membrana rivolta verso il compartimento interno: gli elettroni conseguenti passano dal centro di reazione P680 ai plastochinoni PQ (trasportatori sia di H+ sia di elettroni); i due H+ richiesti per la riduzione di PQ provengono dallo stroma.Gli H+ vengono spinti nel compartimento interno quando gli elettroni passano da PQ al citocromo f, che non trasporta H+ ed è situato sul lato interno della membrana tilacoide. Fra i trasportatori del centro di reazione P700, NADP+ riduttasi, (FAD dipendente), veicola ioni H+ ed elettroni. FAD può veicolare sia H+ sia elettroni ed è rivolto verso lo stroma. La riduzione di FAD a FADH2 utilizza due H+ dello stroma. Gli elettroni e un H+ vengono trasmessi a NADP+; l’altro H+ viene rilasciato nello stroma.
Il trasporto di H+ indotto dalla luce rende lo spazio interno più acido. Il flusso di H+ verso il lato stromale attraverso l’unità F0 determina la sintesi dell’ATP da parte dell’unità F1. Anche NADPH si forma sul lato stromale della membrana. Così ATP e NADPH, prodotti della reazione alla luce della fotosintesi, sono localizzati nella corretta posizione per le successive reazioni n cui la CO2 è convertita in carboidrati.
Gradiente di protoni e flusso non ciclico degli elettroni
Quando gli elettroni fluiscono dal fotosistema II al fotosistema I passano attraverso il complesso del citocromo bf. Questo catalizza il trasferimento degli elettroni dal plastochinone PQ alla plastocianina PC.
La riossidazione del PQH2 ridotto “scarica” nel lume interno dei tilacoidi due protoni che non si legano all’accettore di elettroni (il citocromo f). Si forma così un gradiente protonico per cui l’interno è più acido.
Flusso ciclico degli elettroni fotosintetici

Esiste una via alternativa alla via Z, per gli elettroni che arrivano dal centro di reazione P700 del fotosistema I, che aumenta la versatilità della fotosintesi.L’elettrone ad alto potenziale della ferrodossina Fd può essere trasferito al citocromo bf (invece che a NADP+) e ritornare alla forma ossidata del P700 attraverso la plastocianina PC.Il flusso di elettroni determina solo il trasferimento di H+ nello spazio tilacoide da parte del citocromo. In questo processo viene generato ATP senza la formazione contemporanea di NADP.
Il fotosistema II non partecipa alla fotofosforilazione ciclica; questa avviene quando non vi è più NADP+ per accettare elettroni dalla ferrodossina ridotta.
Catena di trasporto degli elettroni
La catena è costituita da una sequenza di molecole che trasferiscono elettroni da NADH e FADH2 all’ossigeno, con una ben precisa direzione del flusso di elettroni.Le molecole sono: flavine (NAD, FAD), centri ferro-zolfo (Fe-S), ubichinone o coenzima Q, gruppi eme di citocromi, ioni rame, tutti associati a proteine.I trasportatori formano quattro complessi di trasporto di elettroni. Gli elettroni derivano dall’azione di deidrogenasi che utilizzano come coenzimi NAD o FAD.
Gli elettroni vengono trasferiti da NADH a FMN (gruppo prostetico della NADH-ubichinone riduttasi); l’ ubichinone Q si riduce a QH2. Lo stesso avviene a causa della succinato-ubichinone riduttasi.
Gli elettroni passano poi da QH2 alla citocromo riduttasi (che contiene i citocromi b e c1); questa riduce il citocromo c, proteina periferica di membrana, solubile in acqua. Il citocromo ctrasferisce elettroni alla citocromo ossidasi (che contiene citocromo a e a3 e due Cu++).
Fe del gruppo eme e un Cu++ trasferiscono quindi gli elettroni dall’ossidasi all’accettore finale, O2.
Direzione del flusso degli elettroni

Nel diagramma è indicata l’energia libera di alcuni trasportatori di elettroni della catena respiratoria a partire dalla energia libera di NADH presa come zero e la direzione del flusso degli elettroni, sempre in discesa verso l’ossigeno.In tre passaggi della catena si ha una diminuzione notevole di energia libera: è in queste fasi che viene fornita energia per la sintesi di ATP, che permette di conservarla.
E-FMN rappresenta la NADH deidrogenasi ; Q rappresenta l’ubichinone altri simboli corrispondono a diversi citocromi.
Lascia un commento